
Pongámonos en situación: una hormiga abnegada, una cigarra pasota. La primera se prepara a conciencia para la llegada del invierno. La segunda pasa el verano a la bartola poniendo banda sonora al bosque…cri-cri, cri-cri…La llegada del invierno se encarga de poner a cada uno en su sitio: mientras el esfuerzo de la hormiga se ve recompensado, la cigarra languidece de hambre y frío. Seguro que os suena esta fábula clásica de la autoría del griego Esopo, posteriormente recreada por el francés Jean de La Fontaine (1621-1695). Hormiga y cigarra, cigarra y hormiga. Ni Esopo ni La Fontaine dedican una sola línea a las bacterias que coexisten naturalmente con estos dos linajes de insecto, casi seguro porque la primera bacteria no fue avistada hasta 1683, gracias a un revolucionario invento del holandés van Leeuwenhoek: el microscopio.
Efectivamente, todos los metazoos (nosotros incluidos) mantenemos una estrecha relación con el mundo microbiano. Esto es especialmente cierto en el caso de los insectos, muchos de los cuales se benefician de simbiosis estables con bacterias intracelulares (conocidas como endosimbiontes) de las que a menudo depende su propia supervivencia. La adaptación a un estilo de vida intracelular supone grandes cambios en las necesidades de las bacterias endosimbiontes respecto a las de sus parientes de vida libre. En este sentido, la estabilidad y riqueza química que ofrece la célula anfitriona les permite prescindir de un gran número de genes que, desprovistos del efecto ‘protector’ de la selección purificadora (aquella que garantiza el mantenimiento de las funciones esenciales en la naturaleza), acaban por desaparecer del genoma debido a un proceso irreversible de acumulación de mutaciones conocido como “degradación genómica”. Por esta razón, los endosimbiontes de larga duración suelen disponer de genomas reducidos que conservan únicamente aquellas funciones génicas que garantizan el mantenimiento y reproducción de la célula bacteriana así como el suministro de los recursos que necesita el insecto hospedador para sobrevivir en su nicho ecológico. Y a veces ni eso. La degradación genómica es un proceso gradual que amenaza en todo momento con afectar funciones génicas esenciales por lo que, en última instancia, la única forma de evitar la extinción es la incorporación al sitema de un nuevo endosimbionte capaz reemplazar al endosimbionte anterior o, al menos, de aportar las funciones esenciales que éste haya podido perder, dando lugar a un consorcio endosimbionte.
Hormigas y cigarras, como tantos otros insectos, mantienen simbiosis de larga duración con bacterias intracelulares. Las hormigas de la tribu Camponotini viven en simbiosis con la gammaproteobacteria Blochmania floridanus desde hace unos 40 millones de años. Como cabe esperar para endosimbiontes de larga duración, Blochmania presenta un reducido set de genes contenidos en un genoma drásticamente reducido. Ninguno de los genomas conocidos de Blochmania supera las 800 Kilobases (Kbs), lejos de las más de 5.000 Kbs del genoma medio de su pariente de vida libre Escherichia coli. Por su parte, muchas cigarras conviven con un par de bacterias intracelulares: Sulcia muelleri y la Hodgkinia cicadicola, bacteroidetes y alfaproteobacteria respectivamente. Ambas bacterias son endosimbiontes de larga duración pero, mientras que Sulcia ha sufrido un proceso de reducción genómica tradicional, al estilo de Blochmania en hormigas, Hodgkinia muestra un patrón de evolución sorprendente en algunas cigarras llamado lineage splitting (algo que podríamos traducir como “separación de linajes”). Básicamente supone la fragmentación del genoma de Hodgkinia en múltiples subconjuntos de genes (codificados por múltiples subcromosomas) distribuidos irregularmente en la población de células de Hodgkinia que infecta a cada cigarra. Es decir, que cada célula de Hodgkinia contiene una colección de genomas derivados de múltiples eventos de fragmentación del genoma original, como los trozos de un mismo espejo que se hubiera roto varias veces. Los distintos subcromosomas se encuentran en dosis variables y siguen el inevitable camino de la degradación genómica que conduce a la inactivación y pérdida más o menos errática de algunos de sus genes. Una locura. A pesar de esta implosión genómica, todas las células de Hodgkinia contienen al menos los genes necesarios para la expresión génica y la replicación del material genético.
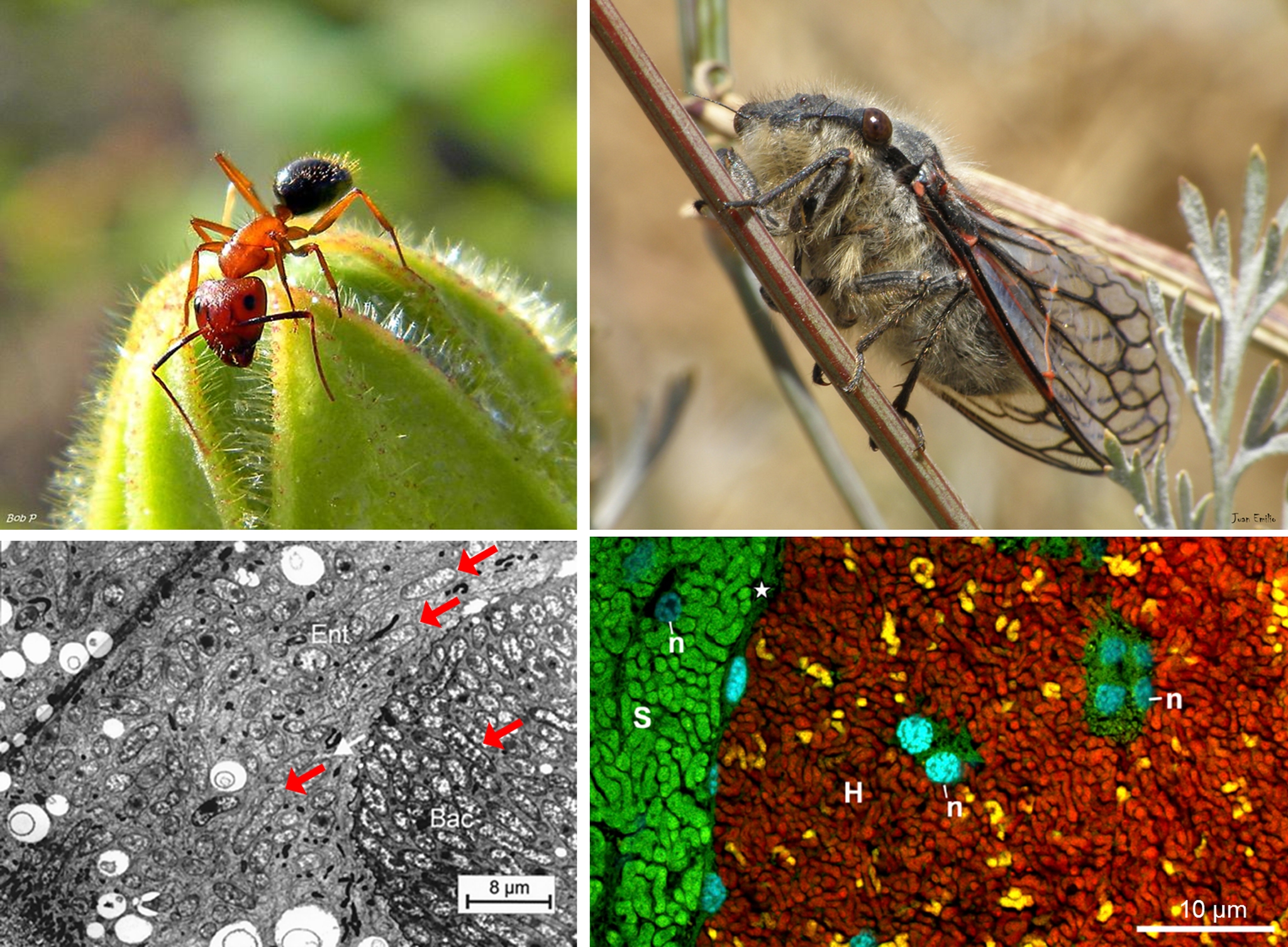
Al estilo de la reducción genómica tradicional, la fragmentación genómica en Hodgkinia parece un proceso progresivo e irreversible. En otras palabras, los trozos del espejo siguen rompiéndose en nuevos trozos; los genes que necesita conservar Hodgkinia siguen repartiéndose en nuevos subcromosomas, lo que dificulta cada vez más que todas las células de esta bacteria sean capaces de heredar un juego completo de genes esenciales en las dosis requeridas. Por el contrario, los genes esenciales se encuentran cada vez más repartidos en la población de Hodgkinia, de modo que ésta deja de ser un simple miembro del consorcio endosimbionte de las cigarras para convertirse en un consorcio endosimbionte en sí misma. Un consorcio dentro de otro, si se quiere. Algo que probablemente disminuye la eficiencia de Hodgkinia como proveedor de nutrientes esenciales en algunos linajes de cigarra.
Curiosamente, los investigadores que han descrito este fenómeno de fragmentación genómica y posterior separación de linajes en Hodgkinia han relacionado la intensidad del proceso con el estilo de vida de las especies de cigarra correspondientes, que pasan la mayor parte de su vida en forma de larvas subterráneas. Así, tasas de mutación elevadas combinadas con ciclos de vida largos facilitarían los patrones de inactivación complementaria que habrían originado el reparto de genes (y subcromosomas) entre las células de la población de Hodgkinia. De este modo, las poblaciones más simples (en términos de implosión genómica) del endosimbionte se han descrito en cigarras periódicas que emergen anualmente, mientras que las más complejas se observan en cigarras de ciclo largo, capaces de emerger solo tras 13-17 años bajo tierra. Algo que nada tiene que ver, por supuesto, con la pereza que les atribuyera Esopo en su fábula. Y sin embargo es la fábula de Esopo lo primero que me viene a la cabeza cuando pienso en cigarras, Hodgkinia y lineage splitting.